

Biology
Southern rock lobster have biological traits that vary across its distribution, including across Tasmanian waters. These biological traits include differences in colouration with red lobsters in the north and east of the state and mottled (green-transparent) in deeper waters in the south west of Tasmania. This colour differentiation is a manifestation of phenotypic plasticity (the ability to change behaviour, morphology and physiology with changes in environmental conditions) that is attributed to differences in habitat and food quality. Faster growth rates are also observed in the north and east of Tasmania and higher quality product in the shallow coastal reefs within those zones. Consequently, spatial management of harvest strategies have been adopted that incorporate variability in biological traits and fleet characteristics with the aim of increasing the yield, value and sustainability of the fishery. For example, regional size limits have been introduced in the north-west that aim to increase egg production and yield.

Lobster sized is measured by the carapace length (CL) from the antennal horns to the end of the carapace.
Length frequency (LF) data is collected by observers on commercial vessels and through scientific sampling. The use of a legal minimum length (LML), which is based on carapace length (CL), has been a consistent management strategy in Tasmanian regulations. LMLs were first approved by the Tasmanian Parliament in the 1885 Act for the Protection of Crayfish and set at 10 inches, but increased in 1890 to 12 inches total length for both sexes. From 1947, legal size was measured by CL and fluctuated between 4 1/6 – 4 1/2 inches until 1966, when a lower size limit for females was introduced. In the 1970s, CL measurements were converted to metric units at 105 mm for females mm and 110 mm for males.
Undersize LF data contributes information on future recruitment in projections, while legal-sized stock is the biomass available for catch and reproduction. We will be adding LF data to the Biology Explorer soon.

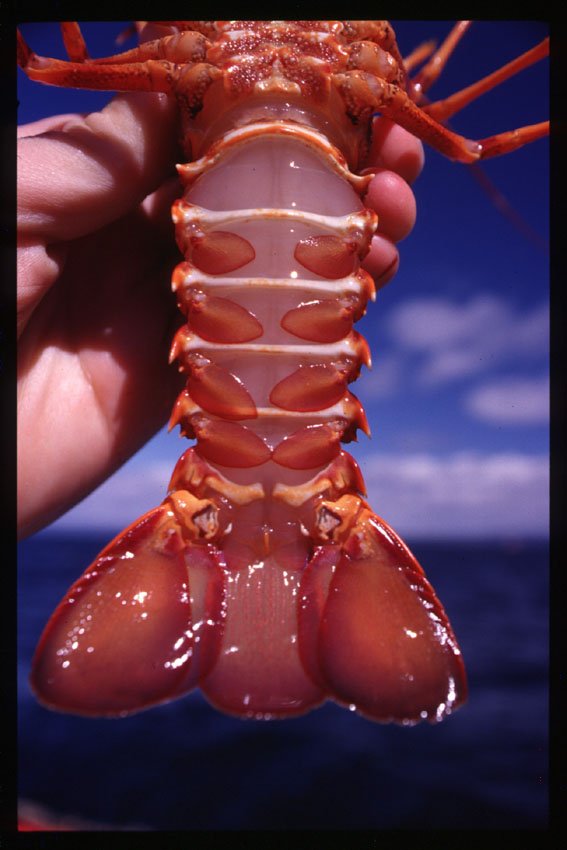
Female (left) and male (right) rock lobster can be identified by the size of the pleopods or swimmerets on the tail. Female rock lobster also have a small nipper claw each rear leg.

Recruitment

Larval Dispersal


